|
Les éléphants de mer
« océanographes »
de l'océan Austral
Christophe Guinet
Chargé de Recherches au CNRS, Coordinateur français
du programme SEaOS, membre du Comité d'experts du Cercle
Polaire
Une vie d’éléphant
de mer
Kerguelen, début du printemps austral, des dizaines de milliers
d’éléphants de mer sont regroupés sur
les immenses plages grises de Kerguelen battues par le vent et le
grésil. Un immense rendez vous annuel où, après
huit mois passés en mer à accumuler des réserves
de graisse, les plus massifs et puissants de tous les pinnipèdes
se retrouvent pour la saison de reproduction.
Les éléphants de mer sont les plus grands des phoques
et, contrairement aux otaries et aux morses, ils ne peuvent pas
utiliser leurs membres postérieurs pour se déplacer
à terre. Ils avancent par une sorte de reptation par ondulations
verticales de la colonne vertébrale, aidée par des
tractions des membres antérieurs. Ce faisant, ils laissent
traîner leurs nageoires à l’arrière du
corps. Les grands mâles (figure 1) de deux à trois
tonnes de muscle et de graisse veillent jalousement sur leur harem
qui regroupe jusqu’à une centaine de femelles, dont
chacune pèse entre 400 et 600 kg.

Figure 1 : couple d'éléphant
de mer austral, Mirounga leonina (CEBC-CNRS / SEaOS)
Malgré son air à moitié assoupi, le mâle
veille attentivement sur ses femelles. À la moindre alerte,
il redresse la tête, exhibant son cou strié de cicatrices,
témoins d’anciens combat de titans. Un nez énorme
qui surmonte une énorme gueule rose se distend sous l’effet
de l’excitation. Ce nez leur vaut le nom d’éléphant,
bien qu’il ne s’agisse aucunement d’une trompe
préhensile.
La période de reproduction est aussi une longue période
de jeûne. Les mâles sont les premiers à arriver
à terre au début du mois de septembre, suivi 2 semaines
plus tard par les premières femelles. Les mâles reproducteurs,
appelés Pachas, vont alors jeûner pendant presque deux
mois, vivant exclusivement sur les réserves de graisse et
de muscle accumulées pendant le long voyage en mer qui a
précédé la saison des amours. Pendant ce jeûne,
ils peuvent perdent jusqu’à 1,5 tonnes qu’ils
dépensent en surveillance, défense du territoire,
combat et parades amoureuses.
Les femelles ne restent à terre qu’un seul mois. Quelques
jours après leur retour à terre, elles donnent naissance
à un unique petit pesant une quarantaine de kilogrammes à
la naissance. Le petit apparaît « tête la première
», revêtu d’une superbe fourrure noire, le Lugano,
qui le protège efficacement du froid (figure 2). Quelques
heures après la naissance, l’allaitement débute
et va se poursuivre pendant trois semaines. Le nouveau né
à la silhouette longiligne va progressivement se transformer
en une petite bonbonne. La totalité du lait qui contient
40 % de matières grasses est produit à partir des
réserves accumulées par sa mère au cours des
huit mois qu’elle vient de passer en mer.

Figure 2 : nouveau-né avec
son Lugano noir (source Wikipedia, B.navez - Kerguelen - 1999)
La masse au sevrage avoisine les 120 kg, néanmoins
de très grandes différences peuvent être observées
entres les petits. Certain ne pèsent qu’une soixantaine
de kilos alors que d’autres peuvent exceptionnellement dépasser
les 200 kg. Or de la masse au sevrage dépendent les chances
de survie du petit. Plus le petit est gros, meilleures sont ses
chances de survivre à ses premières années
en mer. Cette masse de sevrage dépend de facteurs tels que
l’âge ou l’expérience des mères.
Le petit des jeunes femelles, qui n’ont pas totalement achevé
leur croissance et qui manquent d’expérience, est sevré
à une masse corporelle plus faible que celui des femelles
les plus âgées et expérimentées. Mais
les facteurs climatiques et océanographiques déterminent
aussi une grande part des variations de cette masse corporelle du
jeune au sevrage. En effet, ces deux types de facteurs influent
largement sur la quantité de proies que les femelles peuvent
capturer durant les mois qui précèdent leur retour
à terre. Comprendre les déterminants océanographiques
et climatiques de telles variations du succès de pêche
des mères et de la masse au sevrage de leur petit constitue
un enjeu important pour les scientifiques. Il s’agit d’évaluer
les répercussions du développement de nouvelles pêcheries,
potentiellement concurrentes des éléphants de mer
pour les ressources alimentaires, et /ou des changements climatiques
en cours sur les écosystèmes marins polaires, susceptibles
de modifier la disponibilité des proies des éléphants
de mer (effectifs, taille, distribution spatiale…).

Figure 3 : femelles éléphants
de mer prenant leur bain de boue en période de mue (CEBC-CNRS
/ SEaOS)
Quelques jours avant de repartir en mer et de laisser son petit
affronter seul les dangers de la vie, la femelle s’accouple
avec le pacha. Ce dernier veille pour repérer les femelles
en chaleur et profite sans vergogne, l’instant venu, de sa
taille. Utilisant son poids et sa gueule, il immobilise la femelle
réceptive dans un acte qui semble plus tenir d’un combat
de catch que d’un acte d’amour. Comme chez les autres
pinnipèdes, l’œuf fécondé commence
à se diviser plusieurs fois, puis se met en sommeil (diapause),
stoppant le processus de développement. Il ne s’implante
que trois mois plus tard sur la paroi utérine (implantation
retardée) et la gestation à proprement parler peut
alors débuter.
Hormis la reproduction, les éléphants de mer reviennent
à terre pour renouveler leur pelage (figures 3 et 4). En
effet, l’élimination de l’ancien revêtement
de poils et la production d’un nouveau pelage nécessite
une très forte circulation sanguine périphérique
pour permettre la synthèse des éléments constituants
des nouveaux poils. L’eau étant un bien meilleur conducteur
de chaleur que l’air, les déperditions thermiques seraient
trop importantes et consumeraient des réserves énergétiques
indispensables à la reproduction et à la survie de
l’éléphant de mer. Ainsi, d’un point de
vue énergétique, la meilleure stratégie possible
consiste à venir muer à terre lors malgré un
jeûne d’un mois qui est moins coûteux en réserves
de graisse.

Figure 4 : Femelles en mue (CEBC-CNRS
/ SEaOS)
Qui menace l’éléphant de mer ?
La graisse de ce phoque, l’un des combustibles privilégié
pour éclairer au XIX siècle de grandes villes telles
que Londres, était l’objet de toutes les convoitises.
Les éléphants de mer ont payé au cours du XIX
siècle un très lourd tribut aux phoquiers. Leur cousin
du Pacifique Nord a même failli totalement disparaître,
moins d’une dizaine d’individus ayant miraculeusement
survécu à la lance des phoquiers. Dans l’hémisphère
sud, après la surexploitation des éléphants
de mer du XIX siècle, la chasse a été régulée
et les effectifs des principales populations d’éléphants
de mer de l’océan Austral, la Georgie du sud, les îles
Kerguelen et Heard, et l’île Macquarie, se sont progressivement
reconstituées malgré la chasse. À Kerguelen
(figure 5), l’usine phoquière de la SIDAP (Société
Industrielle des Abattoirs Parisiens) exploita jusqu’en 1960
les mâles d’éléphant de mer à partir
de son usine pilote où furent montées des machines
arrivées de France et d'Australie (cuiseur, autoclave, «
égoutteur », centrifugeuse…), ainsi que divers
véhicules utilisés pour la chasse à terre des
éléphants de mer et leur transport jusqu'à
l'usine.


Figure 5 : restes de station de fonte
d'huile d'éléphant de mer à Kerguelen (source
Wikipedia, B. Navez) et dépeçage (SEaOS)
Malgré l’arrêt de la chasse, alors que quelques
populations sont stables, voir même en légère
augmentation, d'autres sont en déclin continu depuis les
années 1960.
Les déclins observés ne semblent pas pouvoir être
imputés à l’exploitation commerciale des éléphants
de mer car, paradoxalement, ce n’est qu’après
la cessation de cette exploitation raisonnée des populations
d’éléphants de mer que les effectifs des populations
de Kerguelen-Heard et de Macquarie ont très sensiblement
diminués au cours des années 1970. En l’espace
de 10 ans, ces populations ont perdu 50 % de leurs effectifs. Une
situation contrastée est observée en Géorgie
du Sud où la population, forte d’une centaine de milliers
de femelles, semble être resté stable pendant cette
période. Le développement des pêcheries australes
est aussi un argument invoqué pour expliquer ce déclin.
L’effet que pourraient avoir eu les pêcheries ne semble
pas résister à l’examen des faits. En effet,
malgré l’absence de pêcheries, les populations
de Crozet, de Marion et des îles Macquarie ont décliné
simultanément et en phase avec celle de Kerguelen où
une pêcherie s’est mise en place. Dans le même
temps, la population de Géorgie du Sud est restée
stable malgré le développement des pêcheries.
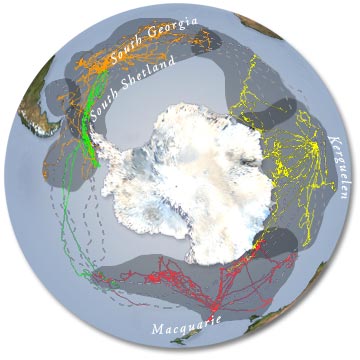
Figure 6 : localisation des quatre
populations d'éléphant de mer austral (SEaOS)
Les raisons de ces variations ne sont pas claires,
mais des changements régionaux d'abondance et de distribution
des proies en relation avec des variations des conditions océanographiques
à grande échelle pourraient être en cause.
Afin de vérifier la plausibilité de ces différentes
hypothèses, il était nécessaire de percer les
secrets de la vie marine des éléphants de mer et de
comprendre les dynamiques très contrastées observées
entre les principales populations dans l’océan Austral
(figure 6). Pour cela, un projet de recherche international, associant
des chercheurs britanniques, australiens et français, a été
lancé en 2004, le SEaOS
(South Elephant seal as Oceanographic Samplers). L’objectif
du programme était d’identifier, au sein de l’océan
Austral, les zones d’alimentation et les domaines océanographiques
exploités par les animaux et de vérifier si le succès
de pêche variait entre populations.
L'éléphant de mer est un prédateur supérieur
qui consomme des quantités considérables de poissons
et de calmars. Il pêche ses proies à de grandes profondeurs
et n'hésite pas, pour les trouver, à entreprendre
d'importantes migrations. Pour mieux connaître les détails
de ces comportements, les éléphants de mer originaires
des trois principaux sites de reproduction ont été
étudiés : l'un en Georgie du sud où la population
a toujours été stable ; un autre à Kerguelen-Heard
où la population s'est stabilisée après une
période de fort déclin ; et un troisième site,
l'Ile Macquarie, où la population diminue toujours.
Pour étudier ces questions, des éléphants
de mer austraux ont été équipés d’une
nouvelle génération de balises Argos, développée
par le Sea Mammal Research Unit
(unité de recherche de l’Université of St Andrews,
Ecosse). Cette nouvelle balise permet le suivi des trajets en mer
des animaux mais aussi de mesurer et transmettre en temps réel
les données de température et de salinité mesurées
au cours de leurs plongées profondes. Les éléphants
de mer deviennent alors de précieux auxiliaires pour l'océanographie
opérationnelle. Ces mesures rendent possible l’étude
de l’hydrologie de l’océan Austral, avec des
résolutions spatiales et temporelles jamais atteintes. Cet
océan demeure encore mal connu, mais il pourrait jouer un
rôle essentiel dans l’évolution attendue du climat
pour les décennies à venir. Par ailleurs, ce programme
d’études nous permet de comprendre comment les variations
des conditions océanographiques affectent le succès
de pêche des éléphants de mer, la condition
des petits issus de la reproduction et les chances de survie des
jeunes après le sevrage.
Entre 2004 et 2006, 85 éléphants de mer ont été
équipés de ces balises à Kerguelen, Macquarie,
en Géorgie du Sud et en Péninsule Antarctique. Cette
étude, initiée par des biologistes marins, a suscité
l’intérêt de plusieurs océanographes dont
le besoin d’une information continue et de qualité
sur l'océan Austral est crucial. Chacune de ces balises fournit,
quotidiennement et en temps quasi-réel, un profil de température
et un profil de salinité à des profondeurs comprises
entre 500 et 1500m, dans une région où les informations
hydrologiques manquent cruellement. Les données ainsi collectées
sont intégrées dans la base de données Coriolis
qui alimente Mercator
en données in situ sur l’observation de l'océan.
Le système Mercator est basé sur une représentation
mathématique (un modèle) de l'océan, contraint
en permanence par des observations satellitaires (hauteur de la
mer, température de surface) et des données in situ
(mesures en mer de température et salinité) qui viennent
"rectifier le tir" en temps réel. Par ailleurs
ces données servent pour des études d’océanographie
physique fondamentale visant à étudier la circulation
des masses d’eau dans l’Océan Austral. Ces données
contribuent aussi à l’observation et au suivi du réchauffement
à moyen terme de cet océan.



Figure 7 : séquence de capture
d'une jeune mâle (CEBC-CNRS / SEaOS)
La pose d’une balise nécessite la capture d’un
éléphant de mer (figure 7), ce qui n’est pas
une mince affaire ! L’animal est puissant et un coup de mâchoire
aurait des conséquences dramatiques. Mais à terre,
l’animal est peu mobile et ne peut que difficilement échapper
aux biologistes. La technique consiste à lui recouvrir la
tête d’un sac qui l’aveugle tout en lui bloquant
la gueule. Ensuite, tout n’est qu’une question de poids
sur l’éléphant qu’il faut immobiliser
le temps nécessaire pour l’ injection de l’anesthésiant.
L’effet de l’anesthésiant est fulgurant et, moins
d’une minute après l’injection, l’éléphant
dort, incapable du moindre mouvement. L’éléphant
de mer assoupi est pesé, mesuré, une prise de sang
est faite et une extrémité d’ongle prélevée
(figure 8).
 
Figure 8 : prise de sang et pesée
de l'animal (CEBC-CNRS /SEaOS)
La balise est collée au moyen d’une résine
à prise rapide sur les poils de haut de la tête de
l’éléphant de mer : c’est la seule partie
de l’animal exposée à l’air lorsqu’il
respire en surface, flottant verticalement dans l’eau (figure
9). Les signaux satellites Argos doivent impérativement être
émis hors de l’eau pour pouvoir être capté
par le satellite. Une fois les manipulations terminées, l’animal
est relâché et la balise, telle un chapeau, le suivra
dans tous ses déplacements. A son retour à terre,
après 8 mois en mer, on re-capture l’animal pour lui
retirer sa balise et effectuer divers prélèvements
et mesures qui seront comparées à celle effectuées
lors de la première capture. Si l’animal n’est
pas re-capturé, la balise tombe naturellement lors de la
mue suivante lorsqu’il revient à terre.
 
Figure 9 : Pose de la balise (CEBC-CNRS
/ SEaOS et SMRU / SEaOS)
Une vie de plongeur d’exception
Autant l’éléphant de mer est mal adapté
à la vie terrestre, autant sa vie marine révèle
les incroyables aptitudes de l’espèce à la vie
aquatique. En mer, il effectue des plongées profondes et
régulières tout au long du trajet. Il passe près
de 90% de son temps sous l'eau, plongeant en moyenne entre 400 et
800 m de profondeur. Mais la plupart des individus peuvent descendre
bien plus profondément, le record étant détenu
par un mâle de Kerguelen à 1 850 m. Dans le monde animal,
seuls les cachalots et certaines baleines à bec semblent
pouvoir concurrencer ces phoques. La durée moyenne des plongées
est de 20 à 30 minutes, à une fréquence de
60 à 80 plongées par jour (figure 10).

Figure 10 : Profils de plongée
d'un individu pendant une semaine d'enregistrement (SEaOS)
Lors de leurs trajets, les éléphants de mer traversent
la plupart des structures frontales et des zones de convergences
péri-antarctiques. Nos éléphants échantillonneurs
collectent ainsi des données à fine échelle,
permettant, par exemple la description des tourbillons et de la
structure des courants dans les principales zones frontales du courant
circum-polaire, ou Convergence Antarctique, mais aussi dans la zone
de la banquise Antarctique. Cette étude montre que la distribution
des éléphants de mer est différente en fonction
du sexe des individus. Les femelles adultes exploitent de façon
privilégiée la zone de bordure des glaces et les tourbillons
des zones frontales polaires. Les mâles concentrent leur activité
de recherche de nourriture principalement sur les plateaux péri-antarctiques
et péri-insulaires.
Ainsi en travaillant simultanément sur les mâles et
les femelles des différentes populations d’éléphants
de mer, nous obtenons des informations océanographiques sur
l’ensemble des régions de l’océan Austral.
Les seules exceptions sont les gyres de la mer de Weddell et de
la mer de Ross que les éléphants de mer semblent consciencieusement
évite (figure 11).

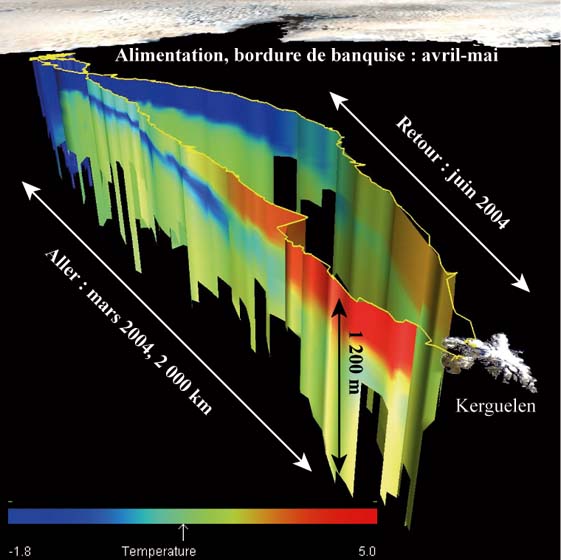
Figure 12 : couverture de l'océan
Austral par le programme SEaOS et exemple de l'enregistrement de
la migration d'un jeune mâle (CEBC-CNRS /SEaOS/programme de
visualisation MANVIS du SMRU)
Les variations du succès de la pêche peuvent être
suivis lors de plongées très particulières,
assimilées à des plongées de repos ou de digestion.
L’animal plonge en profondeur probablement afin d’échapper
à son plus redoutable prédateur, l’orque, qui
est un piètre plongeur. Pendant ces plongées, les
éléphants de mer nagent activement jusqu’à
200 – 300 m de profondeur, puis cessent toute activité
natatoire et se laissent dériver passivement dans la colonne
d’eau. Or le taux de dérive est directement fonction
de l’état d’engraissement de l’animal.
La graisse étant moins dense que l’eau, plus l’éléphant
de mer s’engraisse, plus il a tendance à l’instar
d’une motte de beurre à flotter. Il tend donc à
remonter plus ou moins rapidement vers la surface en fonction de
son état d’engraissement. À l’inverse,
lorsque l’éléphant de mer s’amaigrit,
la proportion de graisse diminue. Devenu plus dense que l’eau
de mer, il tend à couler pendant les phases de dérive.
Ainsi, les variations du taux de dérive des plongées
de repos nous renseignent sur la qualité alimentaire de la
zone visitée par l’éléphant de mer.
Des différences notables dans le choix des zones de pêche
sont observées entre les principales populations d’éléphants
de mer. En Géorgie du Sud, les zones de pêche préférées
sont uniquement situées dans les eaux comprises entre le
front subtropical et le front polaire alors que les eaux antarctiques
ne sont pratiquement pas utilisées par cette population.
À Kerguelen et, dans une moindre mesure, à Macquarie,
ce sont les eaux antarctiques qui offrent les conditions les plus
favorables. Cependant, pour atteindre ces zones favorables et revenir
sur leur site de reproduction, les éléphants de mer
de Kerguelen et de Macquarie doivent traverser des zones particulièrement
défavorables, entre la bordure de la banquise et la zone
frontale polaire, et voyager pratiquement un mois de plus que ne
le font les éléphants de mer de Géorgie du
Sud.
Cette étude confirme la très forte affinité
des éléphants de mer des Iles Kerguelen et Macquarie
avec la glace de mer en hiver, et plusieurs individus passent l’hiver
au cœur de la banquise à parfois plus de 600 km de sa
bordure.
Les diminutions drastiques des effectifs d’éléphant
de mer observées à Kerguelen et à Macquarie
pourraient ainsi être une conséquence directe du changement
de régime climatique et océanographique qui a affecté
l’océan Austral à la fin des années 1960
et souligné par de nombreuses études. Associée
à ce réchauffement, une diminution de près
de 25 % de l’étendue de la banquise a été
décrite et semble avoir eu un impact considérable
sur l’abondance du stock de Krill à la base de la plupart
des réseaux trophiques antarctiques. Les larves de krill
sont particulièrement vulnérables aux variations d’étendue
de la banquise, car les algues épontiques (algues des glaces)
qui se développent sur la face inférieure de la banquise
constituent l’essentiel de leur régime alimentaire.
Or moins la banquise est étendue, moins il y a d’algues
épontiques et par conséquent de krill. Inversement
ces conditions sont favorables au développement d'organismes
gélatineux, les salpes, qui ne peuvent pas être consommées
par la vaste majorité des prédateurs. La diminution
d’abondance du krill a probablement été suivie
par la réduction des effectifs des poissons et des calmars
en bordure du continent austral, les proies dont dépendent
les éléphants de mer de Kerguelen et de Macquarie.
Cette diminution d’abondance des proies, outre un effet direct
sur le taux de survie des adultes, pourrait avoir un effet indirect
par la baisse de la masse corporelle des jeunes au sevrage, réduisant
alors le taux de survie des jeunes.
L'océan Austral, immensité méconnue
L'océan Austral est probablement l'océan le moins
accessible et le moins connu de la planète, en raison de
son isolement géographique et des conditions climatiques
qui y règnent. Or cet océan joue un rôle fondamental
dans les échanges thermiques et de gaz carbonique entre l'océan
et l'atmosphère et, par conséquent, dans la régulation
du climat de notre planète. C'est dans cet océan que
s'effectuent l'essentiel des échanges d'eau et de chaleur
avec les océans voisins (Atlantique, Indien et Pacifique)
et l'atmosphère. On sait maintenant que les variations de
la quantité de chaleur stockée dans les eaux de l'océan
Austral influencent la distribution et l'abondance du krill, et
par conséquent de ses prédateurs : manchots, phoques
ou baleines.
Cette approche est particulièrement appropriée pour
collecter des données hivernales pour l'océan Austral,
qui sont difficiles et extrêmement coûteuses à
obtenir par des méthodes classiques alors que la banquise
recouvre une large surface de la mer. L'emploi de navires océanographiques
conventionnels pour collecter de telles données, outre leur
coût élevé, ne peut pas garantir l'accès
aux zones de glace de mer pendant les mois hivernaux. La seule autre
alternative est l'utilisation de bouées dérivantes
munies de capteurs divers. Cependant cette technologie ne permet
pas une couverture importante des zones de hautes latitudes. Par
ailleurs les bouées dérivantes ne peuvent pas être
dirigées vers des zones présentant un intérêt
particulier alors que le choix d'éléphants mer de
sexe différent permet de cibler des régions particulières.
Les bathythermographes lancés depuis les bateaux de ravitaillement
ou de commerce apportent, elles aussi des informations importantes.
Mais l'obtention de ces données est limitée aux principales
voies de navigation et concerne seulement les profils de température.
Les mouillages fournissent des séries temporelles à
long terme, mais leur nombre est limité compte tenu de leur
coût et des opérations logistiques associées
à leur mise en œuvre. Une révolution dans les
systèmes d'observation est cours, basée sur les satellites
et sur les flotteurs-profileurs (Argos) autonomes qui permettent
d'obtenir pour la première fois des mesures globales pour
l’étude des océans. Mais même ces nouvelles
technologies ne sont pas en mesure d’échantillonner
en routine dans la zone de glace de mer : les mesures par satellite
des températures de surface et de hauteur de la mer sont
impossibles en zone couverte de banquise et les profileurs ne peuvent
pas fonctionner dans ces conditions. Ainsi, pour la zone s’étendant
de 60° S au continent Antarctique et comprise entre 20 °
E et 140 ° E, moins de 300 profils obtenus par ces méthodes
sont répertoriés dans la base de données de
Coriolis Godae. Par contre, en trois ans de programme SEaOS, les
éléphants de mer en ont collecté plus de 5
000. À ce jour, les données « éléphant
de mer » représentent plus 98 % des profils de températures
et salinité obtenus sous la glace de mer (figure 12).
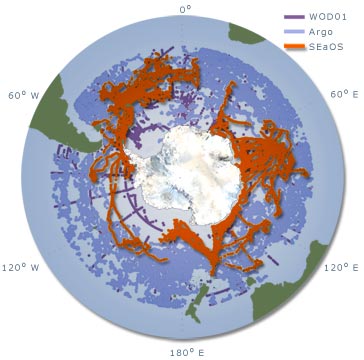 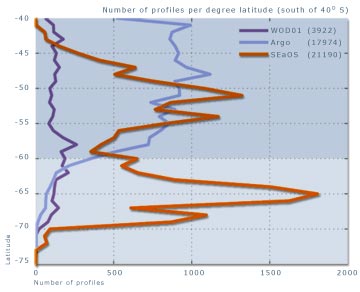
Figure 13 : comparaison du nombre
de relevés et des zones géographiques couvertes par
les différents programmes de relevés océanographiques
dans l'océan Austral (SEaOS)
Une avancée technologique toujours renouvelée
Le développement par les scientifiques et les ingénieurs
du Sea Mammal Research Unit de balises Argos, incorporant dans ces
appareils un capteur CTD miniaturisé (salinité et
température), et permettant l'enregistrement et l'envoi par
satellite des données de plongée et de localisation
d'animaux marins dans leur milieu naturel a grande avancée
technologique rendant possible cette étude est un véritable
progrès pour l’océanographie polaire. L'unité
de base contient une plate-forme de transmission satellite, qui
transmet les données de position via le Service Argos, ainsi
que des capteurs de conductivité (indicateur de la salinité
de l’eau), température et pression (indicateur de la
profondeur de plongé) de haute résolution. Les algorithmes
de compressions des données ont été largement
développés par l’équipe d’océanographie
Physique du Muséun National d’histoire Naturelle. Pour
répondre à l’exigence des océanographes
quant à la qualité des mesures hydrologiques, les
capteurs ont été étalonné avec le concours
du SHOM ( Service Hydrographique
et Océanographique de la Marine française). Puis des
essais en mer ont été réalisés avec
l’aide de l'Insu
(Institut National des Sciences de l’Univers ), et de l’IPEV
(Institut polaire français Paul Emile Victor). Ces test ont
permis de s’assurer de la qualité et de la résolution
des données fournies et d’appliquer le cas échéant
les corrections nécessaires aux données transmises
par chacune des balises CTD.
Dans le cadre des actions menées pendant l’Année
Polaire Internationale, ces efforts de recherches seront approfondis
dans le cadre du programme MEOP
(Marine Mammal Exploration of the Oceans - Pole to Pole). Élargissant
à l’océan Arctique les techniques développées
sur l’éléphant de mer austral, des balises seront
déployées sur les phoques du Groenland pour lequel
tout le monde s’accorde à reconnaître que les
impacts du changement climatique sont plus marqués. Dans
l’océan Austral, nos travaux seront poursuivis pour
préciser les facteurs de vulnérabilité des
populations d‘éléphants de mer aux changements
océanographiques en cours, tout en continuant à utiliser
ces animaux pour observer et mieux décrire l’océan
Austral. L’ensemble des données collectées par
les éléphants de mer équipés à
Kerguelen sont mises à disposition des océanographes.
Désormais, l'obtention de données océanographiques
via des prédateurs marins pour appréhender les conséquences
du réchauffement global s'avère être une approche
très prometteuse pour l’étude des phénomènes
climatique et océanique de grande échelle.
Demain les éléphants de mer permettront de mesurer
les concentrations en phytoplancton de l’océan Austral.
Ce nouveau développement technologique est effectué
avec le soutien du CNES pour obtenir une mesure écologique
particulièrement importante. Dans l’océan Austral,
du fait du brassage permanent exercé par le vent, le maximum
de concentration du phytoplancton n’est pas observé
à la surface, comme c’est le cas dans les zones tempérées
ou tropicales, mais à des profondeurs comprises généralement
entre 30 et 60 m. Par conséquent, la densité du phytoplancton
ne peut pas être mesuré précisément par
satellite qui mesure la concentration de chlorophylle à la
surface des eaux. Ces mesures sont très attendues par les
biologistes marins qui pourront ainsi évaluer beaucoup plus
précisément les quantités de carbone fixées
par le phytoplancton dans le domaine austral et de préciser
le pouvoir de piégeage du gaz carbonique par l'Océan
Austral. D’un point de vue biologique, ces travaux vont nous
permettre de quantifier les variations de productivité de
l’océan Austral en fonction des variations naturelles
du climat et leur conséquence pour un ensemble de prédateurs
marins clefs, dont l’éléphant de mer. Les dernières
informations sont cependant inquiétantes : la masse au sevrage
des éléphants de mer de Kerguelen diminue…
- Christophe Guinet ; Frédéric Bailleul ; Charles
André Bost : Centre d’Etudes Biologiques de Chizé
(CEBC), CNRS, France
- Martin Biuw ; Mike Fedak; Phil Lovell ; Frazer Monks : Sea Mammal
Research Unit (SMRU), University
of St. Andrews, Ecosse
- Jean-Benoît Charrassin ; Young-Hyang Park ; Fabien Roquet
: Equipe "Physique de l'Océan Austral", Département
Milieux et Peuplements Aquatiques, Muséum National d'Histoire
Naturelle, France
- Ian Field ; Mark Hindell; Clive McMahon: Antarctic Wildlife Research
Unit (AWRU),
University of Tasmania, Australie
Retour accueil
Christophe Guinet CEBC-CNRS
/ SEaOS © 2007- Le Cercle Polaire - Tous droits réservés
|


